| CONTROL OF LAMPENFLORA AT WAITOMO CAVES. NEW ZEALAND
KARL JOHNSON
INTRODUCTION The growth of lower plants; algae, mosses and ferns "lampenflora" in the vicinity of lighting fixtures is a well recognised problem in commercial caves. In all caves the naturally occurring levels of nutrients and moisture are sufficient to support plant growth. However, only in commercialised "tourist" caves is the other growth factor, light, provided in sufficient quantity and quality to allow development of coloured patches of autotrophic plants adjacent to the lights. primarily the patches of lampenflora are a visual nuisance in that they obscure and discolour the cave formation but also, the plants produce weak organic acids which in time can erode the calcite and limestone of which the cave formations are made. Therefore it is generally conceded that an attempt should be made to eliminate these organisms. The elimination of the lampenflora involves one or more of the following actions: (a) making tho cave environment locally toxic or inhibatory to plant
(b) the periodic killing and/or removal of the plant populations, or (c) a reduction, to sub-growth levels, of the quantity and/or quality
of
This study of the lampenflora in the Waitomo Caves and the possible methods for control of plant growth in the caves was started because it was recognised that after many years of operating as tourist caves, the three Waitomo caves were undergoing changes in their natural environment, including the growth of substantial lampenflora populations which could lead to permanent deterioration of the caves. The methods investigated for control of lampenflora in commercial caves outside of New Zealand have been discussed by Kermode 1975). The two most successful methods reported have used either steam and/or hot water to both kill and remove the lampenflora (Aley. 1972) or a formaldehyde solution to kill and bleach the lampenflora (Leferur and Laporte. 1969). The research reported here was undertaken to establish what possible methods of lampenflora control could be used in the Waitomo Caves. Primary consideration in the methods tested was for their: ecological acceptability particularly with regard to other cave organisms such as the Glow-worms, practicality with regard to continuing public access and safety, and immediate applicability such that lampenflora control could begin without delay. Therefore, the work reported has centred around two main areas: the use of phytotoxic chemicals to kill and retard regrowth of existing lampenflora populations, and the possible control of cave lighting to retard or prevent population regrowth after chemical cleaning. WAITOMO LAMPENFLORA The plants found in the Waitomo Caves referred to as "Lampenflora" belong to four major groups: algae, mosses, ferns and fungi (See Table 1). These groups, while occupying a similar habitat exhibit two and possibly three modes of energy uptake. The mosses and ferns are obligate auto- trophs requiring light energy for photosynthesis and growth. The algae, both green and blue-green, are basically autotrophic but some species may also be myxotrophs (Kermode 1975, p.339) and thus may be able to utilize the energy from both light and organic molecules for growth. The fungi are obligate heterotrophs obtaining all of their energy requirements from the breakdown of organic materials and therefore do not directly require light for growth. The growth of autotrophic plants in any environment is dependent on
the availability of adequate levels of three essential factors: light.
moisture and nutrients.
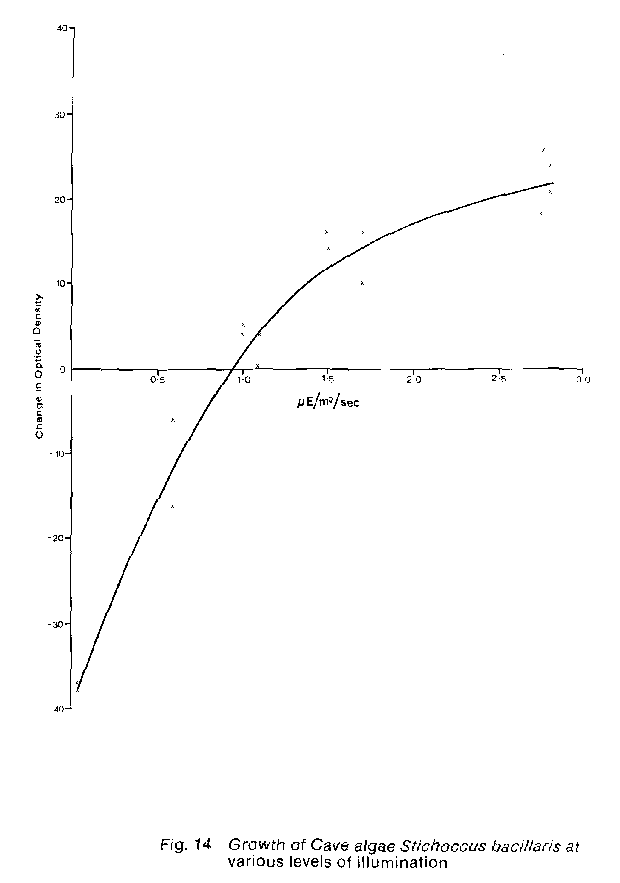
Generally adequate levels of water and nutrients are available in the environment of all caves, commercial or wild.Thus* species isolated and grown in laboratory culture. only the introduction of artificial lighting in commercial caves such as the Waitomo caves is necessary to provide an ideal environment for the growth of autotrophic plants. Unlike the autotrophic plants, the fungi do occur in both wild and commercial caves and do not themselves require light. However, it is only in commercial caves that organic matter is produced by autotrophic plants in sufficient concentrations to permit significant development of fungal populations. Within the three Waitomo tourist caves it appears that the close interaction of the three essential growth factors controls the growth, development and specific composition of the lampenflora. Light - The restricted occurrence of lampenflora to areas directly lit by the fixed lamp housings in the caves clearly shows that light is the primary controlling factor for lampenflora development. The extent of lampenflora growth around any one lamp housing depends on the number of bulbs in that housing and on the distance of that housing from a suitable substrate. Light intensity measurements made in the Glow-worm and Ruakuri caves suggest that the minimum light levels required for continued growth of the different autotrophic lampenflora organisms are: green and blue- green algae 0.1 to 1.0u E /m2 /sec ( 10 to 50 lux) , mosses 1.0 to 3.5u E / m2/sec ( 50 to 180 lux), and ferns 5.0u E/m 2/sec ( 250 lux). A laboratory experiment was undertaken to try and evaluate the growth of one species of cave algae, Stichococcus bacillaris, at low levels of illumination. This species was grown in an enriched soil extract liquid medium to 150ml flasks, at 20 C under 24 hour illumination from coolwhite fluorescent tubes on an orbital shaker table operating at 100 oscillations per minute. Pairs of flasks were equally shaded such that each pair received a different level of illumination ranging from 0.05 to 2.80u E / m 2 /sec. The growth of the algal population in each flask was monitored at weekly intervals for four weeks by measuring the optical density of chlorophyll in the living population. The optical density was measured at 678nm as suggested by Sorokiu (1973 pp. 342-3) on a Beckman Spectronic 700 spectrophotometer. The results of this experiment, (Fig. 14) indicate that under the experimental temperature and light conditions a population of A. bacillaris requires more than 0.9u E/mW / Sec illumination for growth. This result is much too high when compared to the lower light intensities known to permit growth in cave populations. Two reasons for this difference in the required illumination for growth between the experimental and it situ populations are suggested: (I) within the laboratory cultures a previously unknown change in growth morphology artificially depressed the indicated growth at low light levels. At light intensities l.5u E/m 2/ sec S. bacillaris grew as a dispersed population of single cells evenly distributed through the medium. However, at light intensities of l.5u E/m2 /sec the algal cells showed an increasing tendency to clump. Thus, the measured optical density of chlorophyll in the low light cultures was inaccurate because of the uneven distribution of the algal cells. (2) it has been shown by Aruga (1965, p. 284) that in natural algal populations a decrease in temperature results in a greater depression of respiration than photosynthesis so that population growth can occur at low light levels at low temperatures. since it was not possible to duplicate the 10-12'c temperatures found in the caves a test of the temperature effect was not made. However, it seems likely that at cave temperatures growth could well occur at much lower levels of illumination than was indicated in the experiment. Observations suggest that established lampenflora populations, can survive long periods of much lower levels of illumination than those cited above. During the course of an experiment on the development of lampenflora under different colours of light (discussed later in this report) a lamp failed after the growth of a lampenflora population. This lamp was not repaired for five months, and after this period of essentially zero illumination the associated lampenflora appeared to have suffered little or no deterioration. There are two possible mechanisms by which long periods of darkness are tolerated. The first, as mentioned by Kermode (1975, p. 339) is that certain algae can "... change from autotrophy to myxotrophy and finally to heterotrophy ...". The second possible mechanism is that the plants can undergo physiological changes (e.g. a much reduced rate of dark respiration) which permit survival during long periods of low light or darkness. Moisture - the normally discontinuous or irregular distribution of the lampenflora around any one lamp housing is predominantly caused by differences in the available moisture in or on the substrate. These differences arise through the interaction of two variables; the nature of the substrate and the regularity of the moisture source. The substrates available in the Waitomo caves are of three types: mud/clay, porous limestone/leucite and hard surfaced calcite. The moisture source c an also be characterised as either a continuous or a periodic supply of percolating ground water. Observations of discrete populations of lampenflora clearly shows that the distribution of different plant groups is at least partly dependent on the various combinations of moisture regime and substrate type. The ferns, because of their extensive root system and upright habit are restricted to moist mud/clay or extremely soft porous limestone substrates. The mosses, which require a large amount of moisture are usually found on mud/clay or porous limestone/calcite substrates. Both of these substrates have a high moisture holding capacity and thus there is an adequate supply of water for plant growth even if the source of ground water is periodic. On hard substrates mosses only occur where there is an essentially continuous supply of ground water. The algae appear able to withstand much greater fluctuations in available moisture than the mosses and ferns. They may occur on any substrate regardless of whether there is any visible moisture present. This capability is probably due to both a physiological adaptation to dehydration as well as a positional advantage resulting from growing as a thin layer directly on the substrate and thus being able to utilise all available moisture. close adherence to the substrate also places the algae within the calm boundary layer adjacent to the substrate surface thus protecting them from drying air currents. The proximity of the lamp housing can also effect the moisture available to the lampenflora. Heat from the lamps can dry the adjacent substrate surface if the supply of moisture is low or sporadic. This drying results in a barren patch nearest the lamp. surrounded by a healthy lampenflora. Nutrients - the ultimate source of almost all the nutrients necessary for autotrophic plant growth is percolating ground water. In its passage, first through the soil and then the limestone, this water picks up soluble inorganic nutrients which are thereby introduced into the cave. In a non commercial cave there are two possible fates for these nutrients, either they are flushed through the cave by continued water movement or they are concentrated by the evaporation of the carrying water. In a commercial cave either of the above may occur. or the nutrients may be taken up by the lampenflora and thus held, at least temporarily within the cave. Observations of distribution and luxuriance of lampenflora growth in the three Waitomo Caves suggest that nutrients bound to particles of clay are a major source of nutrients for the lampenflora. This is suggested as part of the reason for the dramatic growth of lampenflora seen on mud or clay substrates where they occur naturally or where this type of substrate has been used to camouflage electrical wiring. There is a third source of nutrients for the lampenflora which while not a major source does lead to a further accumulation of nutrients within a commercial cave. This source involves the release of nutrients contained in the organic matter produced by the autotrophic organisms, through the action of the heterotrophic fungi followed by uptake again. Thus, within the lampenflora population itself there may be a cycle of nutrient utilisation, release and utilisation by the autotrophic and heterotrophic organisms. Laboratory Cultures Laboratory cultures of selected lampenflora species were established to serve as test organisms during this study (see Table 1). These species were isolated by the streak plating technique (Hoshaw and Rosowski 1973 p. 58) from samples of lampenflora populations collected in the Glow-worm and Ruakuri Caves. The cultures were grown at 20'c and a 16:8 photoperiod on a soil extract-agar medium (Nichols 1973 p. 19) enriched with 9mg KH2 PO4 and 80mg KNO3 per litre. LAMPENFLORA CONTROL INTRODUCTION The control of lampenflora in any established commercial cave like the three Waitomo Caves, involves two operations: one, the elimination of existing plant populations; and two, the prevention or retardation of their regrowth. A number of ways of accomplishing these two objectives have been suggested by overseas experience and by previous experiments carried out in the Waitomo Glow-worm Cave (Kermode 1975, p. 340). These methods of lampenflora control fall into three main categories: Physical control, chemical control, and Environmental control; each of these are discussed separately below. The methods selected for trial in this study were first evaluated on the basis of ecological acceptability and safety, together with consideration of cost and suitability for general application. Physical control - The general Use of any harsh physical or corrosive methods of lampenflora control is not advocated. This recommendation is based on the deleterious effect these materials and techniques would have on the soft speleothem formation in the Glow-worm Cave and the long-term effect on surfaces in the other caves. The methods included in this category are the use of: stiff brushes, abrasive cleansers, high pressure water and strong acids. There may be a case of using those techniques for a single or extremely infrequent application in specific areas of the caves, but they are claimed inappropriate for general or repeated use. The single exception to this might be the general use of high pressure water once to remove dead plant material and clean the cave surfaces after treatment by phytotoxic chemicals. Chemical control - Early in this study it was realised that chemical control of lampenflora offered an attractive solution to the problem. However, the choice of phytotoxic material used required consideration of many factors which eliminated the possible use of certain groups of compounds. The personal safety of the cave guides, the visiting public and both people and livestock downstream from the caves particularly the Glow-worm and Ruakuri Caves, eliminated the possible Use of two large groups of phytotoxic chemicals. These groups were, those contain highly toxic materials such as arsenic, cyanide, lead or mercury; and, materials of moderate toxicity, which are not quickly degraded which might therefore accumulate within the caves such as chlorinated hydrocarbons and other carbonate compounds. In considering the effect of any chemical on glow-worms, it is not enough to allow only for possible direct contact. It is also necessary to evaluate possible effects of exposure to volatile products or by-products from the phytotoxic chemicals used. Thus, highly aromatic compounds such as formalin or acrolein were not considered for use. A number of other phyotoxic agents are also eliminated strictly on the basis that insufficient information was available to assess their potential hazard in the Waitomo caves environment. Consequently only five potential phytotoxic substances were chosen for further evaluation by insitu trails in the caves. These trails confirmed that a practical and safe method of eliminating and controlling lampenflora growth is possible using chemical phytotoxins. Environmental control - Theoretically the ideal way to kill and /or prevent the growth of lampenflora would be to temporarily or permanently alter the environment in which it lives such that growth is not possible. Three methods of lampenflora control have been suggested which accomplish this objective: (1) temporally exposing the lampenflora to lethal temperatures, e.g. 70'c+ by the use of low pressure hot water or steam (Aley 1972. p. 33); (2) periodically exposing the lampenflora to lethal intensities and wavelengths of ultra-violet light, e.g. 150 to 250 u W/cm2 at 200 to 300 nm (Kermode 1975, p. 340); or (3) altering the quality or quantity of light used for illumination in the caves to a condition which will not support plant growth. The hot water/steam technique for lampenflora removal and cave cleaning has been given extensive trials in the Glow-worm Cave (Kermode 1975, p. 341-2). From these trials and overseas experience (Aley 1972) this method has been shown to be effective, and it has the advantages of highly localised effect and a lack of any toxic residues. However, there are also a number of disadvantages which must be considered. In the Glow-worm cave the soft condition of some spelothem surfaces means that even low pressure steam or hot water will erode these surfaces somewhat and repeated applications could result in serious damage (Kermode 1975. p. 341). Also, this method of treatment is not easily undertaken, as the equipment needed is bulky and requires ready access to a steady supply of water and energy. Finally, this method is not fast, as Aley (1972, p. 34) reports that ten minutes are necessary to treat one square metre of cave surfaces. these disadvantages are not insurmountable with regard to the Glow-worm Cave which is fairly small and has piped water available. However, for this method of lampenflora control to be used in either Ruakuri or Aranni caves a considerable amount of time and manpower would be required just to move the equipment and supply it with water. The use of steam or hot water for initial lampenflora control may be warranted, if a general cave cleaning programme is undertaken. But, the repeated use of this method for long-term lampenflora control is considered unsatisfactory tar the Waitomo Caves. The use of ultra-violet light to kill and prevent regrowth of lampenflora has been discussed by Kermode (1975. p. 340). From this an experiment to test the gross effect of short wave ultra-violet light on lampenflora was undertaken as part of this study. The results of this experiment indicates that this method of control is impractical. particularly as reported by Kermode (loc. cit.) ultra-violet radiation, from a 30 W lamp. is effective only to a distance of 50 to 70 cm from the light source while lampenflora is often found growing in excess of 3m from existing lamp housings. Therefore to totally control lampenflora by this method would require lamps of about 400 W output positioned with existing lamp housings or multiple low power lamps positioned closer to the lampenflora substrate. In either case the costs alone would make such a method of lampenflora control impractical. Altering the quality or quantity of light used for illumination in the caves to control lampenflora can be accomplished in two ways; one, to use colours of light that cannot be utilised by plants for photosynthesis; or two, by reducing the intensity or duration of illumination below the level required for lampenflora growth. The first of these alternatives was tested by experimentation in the Glow-worm Cave, the second has been evaluated by an eleven month study of present lighting practices within the three Waitomo Caves. From these studies it does not appear that control through the use of non-photosynthetically active light is feasible, and, that total control of lampenflora through reducing light intensity and/or duration cannot be accomplished throughout the caves because of the minimum light levels needed for visitor safety and enjoyment of the caves. However, the adoption of the recommendation on lighting practice made later in this report concerning the control of cave lighting would greatly reduce the amount of lampenflora growth, and therefore the need for frequent applications of other control methods. Chemical Control On the basis of criteria mentioned elsewhere the five phytotoxic chemical
compounds chosen for use in the Waitomo Caves were: Diquat (6, 7 -
The initial tests of these compounds in the caves consisted of spraying separate 10cm square areas of established lampenflora with each of these chemicals. The results of these tests showed that the two photosynthetic herbicides had no apparent effect. while all three of the inorganic chemical compounds clearly reduced or eliminated the lampenflora in the test squares (see Table 2). Between the inorganic chemicals the major difference seen was in the speed of phytotoxic effect. The two hypochlorite treatments showed obvious death of lampenflora within 12 hours, whereas the effect of the Borax treatment was not apparent until about one month after application. The lack of visible effect of the photosynthetic herbicides Diquat and Diuron was unexpected. subsequent study of reports on the way these compounds are thought to act suggest that their failure was caused by a combination of the low light levels under which the plants live and the rate of degradation of the herbicidal compounds. In both cases the toxic action of these herbicides is the result of an interruption of photosynthesis and the formation of a secondary toxic in the plant treated (Ashton & crafts 1973 pp. 195-6, 386-7). It is suspected that, because of the low light levels in the caves, the toxic action of the herbicide was too slow compared to its rate of degradation and therefore toxicity was never reached lethal levels. In comparison to the photosynthetic herbicides the three chemical herbicides tested are toxic to plants when applied and do not require light to be effective. The two hypochlorite compounds are quick acting penetrate poisons. Therefore, when sufficient hypochlorite is applied, plant death occurs soon after and is indicated by bleaching. Since both of these compounds degrade rapidly, particularly when in contact with organic matter, their effectiveness is dependent on the concentration of hypochlorite in the solution applied and on the density of this plant material to be treated. The noted difference in effectiveness of the two hypochlorite compounds is probably due to deterioration of the Sodium hypochlorite used. This compound is available only in aqueous solutions and in this form is unstable and thus deteriorates with time unless it is stored in the dark under refrigeration. The killing action of Sodium tetraborate results from the excessive uptake of boron by the treated plants. In very small quantities, as it occurs naturally, boron is an essential nutrient for plants. However, when a high concentration of this element is present the plants will take up a toxic quantity. Therefore. when this compound is used as an herbicide it is applied such that a toxic concentration is present in the plant's environment over a sufficient period for lethal uptake to occur.
Laboratory experiments were conducted to find out what concentration
of sodium tetraborate was necessary to totally inhibit growth of lampenflora
organisms. In these experiments each lampenflora species which had
been established in unispecies culture was inoculated on a soil-extract-agar
medium containing from 0 to 800ppm (0 to 800 mg/l) sodium tetraborate.
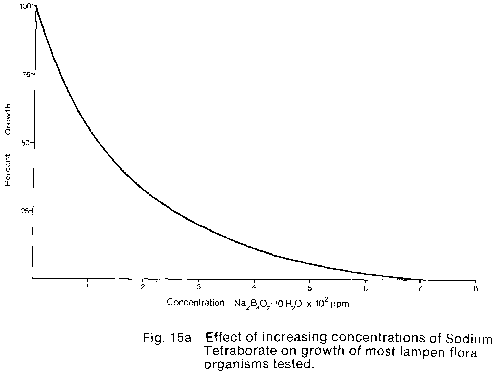
Most of the orqanism tested showed a high initial inhibition of growth
with a decreasing rate of inhibition with increasing concentration. (Fig.
15a) . Total inhibition in this group occurred between 500 and 700ppm
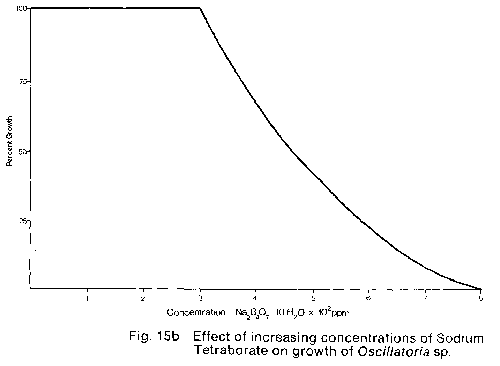
Sodium tetraborate. In contrast. the Oscillatoria sp. tested was
resistant to concentrations up to 300 then showed rapidly decreasing growth
with increasing concentration with total inhibition occurring at
Based on the results of the small scale trial and consideration of the ecological impact of the chemicals involved calcium hypochlorite was chosen for large scale cave trials. Calcium hypochlorite was selected in preference to sodium tetraborate for two reasons: (1) Sodium tetraborate is a non-degradable compound which is dissipated only by leaching with water, while Calcium hypochlorite is degraded within 5 min to 12 hours after application (A.W. Haugley pers. Comm.). Thus, the use of sodium tetraborate over a period of low rainfall and therefore ground water percolation could result in a build-up of the chemical within the caves. This build-up could then be released over a short period, after substantial rainfall, which could result in uncontrolled toxic levels of Boron in the caves and in the Waitomo stream. (2) sodium tetraborate is slow acting requiring 2 to 4 weeks for visible results while Calcium hypochlorite kills and bleaches the plant material within 30 min to 2 hours. Therefore, since two and sometimes three applications of phytotoxic chemicals are needed for a 100% kill, the use of Sodium tetraborate could require 6 to 12 weeks before a given lampenflora population was dead. whereas using calcium hypochlorite the bleaching action of this compound gives visible proof of a complete kill within 6 hours. In the large scale trials of calcium hypochlorite carried out in the Glow-worm and Ruakuri Caves it was found that the concentration of hypochlorite needed for effective control depended on the composition and density of the lampenflora to be treated. where the lampenflora consisted only of a thin covering of algae, then a 2% solution* was strong enough to clear the surface. If however, the lampenflora was made up of a dense matt of mosses and algae then a 4% solution* was necessary. The only major problem noted concerning the use of Calcium hypochlorite
is the strong chlorine smell evident immediately after treatment.
The actual concentration is very low but human sensitivity to this chemical
is such that decided irritation is noted. This problem is only serious
in unventilated areas of the caves, e.g. the Organ Loft of the Glow-worm
Cave. However, 12 hours after treating the Organ Loft area with four
litres of 4% hypochlorite only traces of chlorine could be detected, and
after 36 hours the smell was undetectable (as judged from Cave Guides comment).
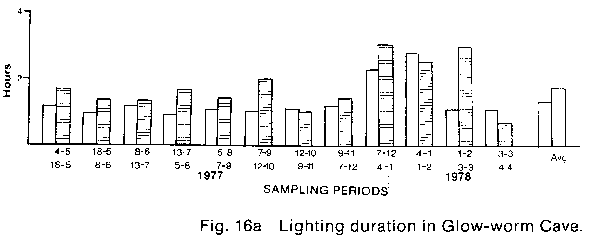
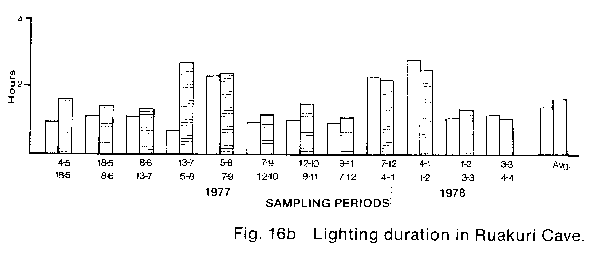
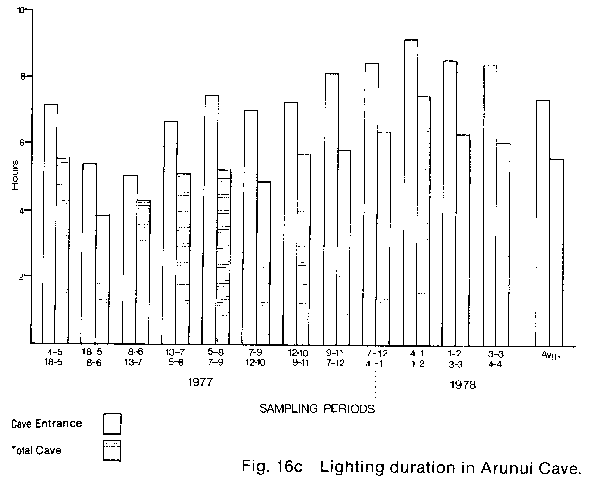
* The percentage hypochlorite used is a weight to volume ratio of commercial granular HTH brand calcium hypochlorite in water. The commercial product actually contains 70% available hypochlorite therefore the true percentage of hypochlorite in the two solutions tested was 1.4% and 2.8% respectively. Lighting control The potential control of lampenflora by modification of present lighting practice in the Waitomo caves involves either the reduction of light quantity and/or the alteration of light quality. If it is assumed that lampenflora growth is proportional to the duration of lighting, then any reduction of lighting duration will result in a reduction of lampen- flora growth. Also it is known that blue light at wavelengths of 350 to 530nm and red light at 600 to 700 nm (Bickford & Dunn 1972) are the colours chiefly responsible for photosynthesis, therefore the reduction of lighting in these wavelengths may also reduce lampenflora growth. To establish what reduction in duration of lighting was possible in the caves, an eleven month study of present lighting duration was made (Fig.16). AMF Vennerette time clocks were modified to function as cumulative hour recorders and were connected to existing lighting circuits at numerous locations throughout the three caves. The locations of the timers were selected such that whenever possible each lighting switch box controlled at least one timer. These timers were then read at two to five week intervals over the eleven months from 4 May 1977 to 4 April 1978. The readings were then converted to average daily hours of lighting (see Appendix 1). The resulting data showed a considerable variation in apparent lighting efficiency between the Glow-worm Cave and Ruakuri and Aranni caves. In the Glow-worm Cave the daily average hours of lighting follows the expected pattern of high values at the cave on entrance, and at the entrance to the Cathedral and low values at the Organ Loft and near the Demonstration chamber. This pattern is shown graphically in Fig. 16A where the cave entrance average (clear bar) is compared with the total cave average (shaded bar) for each sample period. This graph clearly shows the difference resulting from the reduced lighting periods in the Organ Loft and cathedral. Also clearly indicated is the variation in lighting over the year resulting from seasonal differences in tourist numbers and therefore the number of tours going through the cave each day. The eleven month data is summarised in the final pair of bars on the graph which shows the eleven month averages. From the eleven month averages it was calculated that total cave lighting was 24% less than entrance lighting. The results of an identical analysis of the timer data from Ruakuri Fig. 16B and Arunui caves Fig. 16c shows a considerably different result. In both of these caves it is apparent that the lights are left on for considerable periods when no tours are in these caves. In fact in both caves there were only six out of twenty-four samples periods when the total cave lighting average was less than the entrance average. The eleven month averages show that the total cave lighting exceeds the entrance lighting by 23% in Ruakuri Cave and 14% Aranui cave. These percentages are particularly significant because both of these caves are essentially in-out eaves which do not have tourist parties passing one point in the cave repeatedly during a tour as happens at the cathedral entrance in the Glow-worm Cave. From these results it is obvious that a significant reduction of the quantity of light available for plant growth can be achieved in Ruakuri and Arunui caves. In the Glow-worm Cave no such obvious excess lighting is apparent. At the beginning of this study an experiment was set up by Dr. P.1. Dromgoole (Botany Department, University of Auckland) in the Blanket Chamber of the Glow-worm Cave to test the effect of different coloured commercially available lamps, on the development and growth of lampen- flora. In general the results of this experiment were negative with regard to providing a means of lampenflora control, but the results demonstrate some interesting features of the development of lampen flora in the caves. In all cases lampenflora development was apparent on substrates adjacent to the lamps after a comparatively short period of exposure. Different colours of light were noted to have a noticeable effect on the composition of the lampenflora and in a number of cases the lampenflora that developed was composed of plant species previously unknown in the Waitomo caves. Also, from this experiment it is apparent that even if a suitable non- photosynthetically active light source was available it could prove to be unacceptable on aesthetic grounds. The use of coloured lamps greatly alters human perception of the physical formation in the caves and eliminates many of the colour contrasts in the cave formations which gives them much of their unique character. RECOMMENDATIONS The results of this study suggest that management recommendations in three areas are necessary to safely eliminate the present lampenflora and to prevent its regrowth. First; spraying the existing lampenflora populations with Calcium hypochlorite to kill and bleach the plants. Second; to modify the present method of cave lighting control to reduce the possibility of lamps being left on for long periods when they are not needed. Third; to institute a continuing programme of cave cleaning and maintenance to limit the possible sites for lampenflora development. Calcium hypochlorite - the ability of Calcium hypochlorite to safely kill living populations of lampenflora has been shown earlier in the report. However, there are two aspects of the use of this compound which must be considered when planning a large scale. long-term programme of lampen- flora control; its limited phytotoxic effect and its unpleasant odour. With regard to toxicity one of the advantages of using Calcium hypochlorite is the hi9h rate at which it breaks down into non-toxic components. However, this high rate of breakdown means that redevelopment of the lampenflora may begin very shortly after the initial treatment, therefore periodic retreatment will be necessary. Unfortunately the one year limit of this study did not permit a further evaluation of this aspect of control. However, theoretical analysis of the problem suggests that even with more time for experimentation no precise timetable for re-treatment could be produced. It is apparent that the rate of lampenflora growth, and therefore regrowth. varies widely both within and between the three Waitomo Caves. This variation, resulting from different levels of light. moisture, and nutrients in the caves, means that no prescribed frequency of treatment will serve all situations. Also, the effect on regrowth of implementing the recommendations on lighting control discussed later, would significantly alter any timetable based on the present situation in the caves. In view of the present impossibility to predict the time interval between treatments it is suggested that after the initial treatment to kill the existing lampenflora. repeat treatments be applied, as necessary. based on three-monthly inspections of the caves. Further, if this policy is carried out and re-spraying is done as soon as regrowth is noticed, it is possible that all re-spraying could be done with hypochlorite solutions of 2 or even 1 per cent. Every attempt should be made to Use the lowest concentration possible as infrequently as possible. This suggestion is not based on any apparent hazards connected with using Calcium hypochlorite. It is the result of a general appreciation that any "foreign" material introduced into the cave environment may have long-term effects that are not immediately apparent in the short term study used here. The release of chlorine gas during the breakdown of the Calcium hypochlorite requires that spraying be done at appropriate times, e.g. when it is known that no tourist parties will be in the caves for a number of hours. Also, it would be best to spray only limited areas at any one time so that natural cave ventilation can dissipate the chlorine within a reasonable time. only in Arunui cave could there be any need to restrict tourist access after spraying with hypochlorite. This cave is both very deep and very poorly ventilated compared with either the Glow-worm or Ruakuri Caves. Lighting Modifications - The present problem of excessive lighting in the Waitomo Caves is caused by the cave guides sometimes neglecting to switch off lamps as a tourist party leaves a cave or an area in a cave. The easiest solution to this problem would appear to be the replacement of the multiple switches on any one box with a single switch mounted visibly on the front of the box, which controls all the lighting circuits in that box by activating a multiple relay. A second modification of present light switching which could further
reduce the total hours of lighting in specific areas of all three caves
is also possible. Cave lighting serves two distinct functions:
(i) general illumination to permit safe passage through the cave;
(ii) specific illumination to highlight special formations.
In a number of places in the caves these two functions are controlled by
separate switches which are all turned on and off at the same time.
It is necessary that the general lighting be switched on and left on while
the tourist party is in the area controlled by any particular switch box.
However, in many cases it would be possible to control the highlight illumination
by momentary
Cave Cleaning and Maintenance - As was discussed at the beginning of this report, two sources of nutrients for lampenflora organisms are mud and decaying organic matter. In many areas of all three caves those two nutrient sources occur naturally and are a part of the "natural cave environment". However, in other areas accumulations of mud and organic matter have resulted from past cave management practices. A progressive programme of cave cleaning is recommended. Costs - The costs of implementing the above recommendations should not be excessive, particularly in view of the potential saving in electricity cost that will be made after the modification of light switching. Based on the present daily average lighting of Ruakuri and Aranui Caves reported earlier, and assuming a reduction in average total cave lighting similar to that seen in the Glow-worm Cave it should be possible to reduce electricity consumption by 7,320.5 kwh/yr in Ruakuri Cave and 7,880.9 kwh/yr in Arunui Cave. These savings would represent 152/yr for each one cent of the current cost of one kwh of electricity. APPENDIX 1
GLOW-WORM CAVE
ARANUI CAVE
RUAKURI CAVE
REFERENCES ALEY, T. 1972. Control of unwanted plant growth in electrically lighted caves. Caves and Karst. 14(5): 33-35. ARUGA, Y. 1965. Ecological studies of photosynthesis and matter production of phytoplankton I. seasonal changes in photosynthesis of natural phytoplankton. Botanical Magazine. (Tokyo) 78: 280-288. ASHTON, F.M. and CRAFTS, A.S. 1973. "Mode of Action of Herbicides. John Wiley and sons, New York. 504 pp. BICKFORD, E.D. and DUNN, 8. 1972, "Lighting for Plant Growth". Kent state university Press. ix-221 pp. HOSHAW. R.W. and ROSOWSKI, J.R. 1973. Methods for microscopic algae. in Stein, J.R. 1973. 53-68. KERMODE, L. 1975. Glow-worm Cave. Waitomo. Conservation study. New Zealand speleological Bulletin 5(91): 329-344. LEFEYRE, M. and LAPORTE, 0.8. 1969. The "Maladie Verte" of Lascaux, diagnosis and treatment. studies in Speleology. 2(1): 35-44. NICHOLS, H.W. 1973. Growth media-freshwater. In Stein, J.R. 1973. 7-24. SOROKIN. C. 1973. Dry weight, packed cell volume and optical density. In Stein, J.R. 1973. 321-344. STEIN, J.R. (ed) 1973. "Handbook of Physiological Methods: Culture Methods and Growth Measurements" Cambridge university press. Cambridge. xii - 48B pp. WILLIAMS, p. (ed) 1974. "Report on the conservation of Waitomo Caves". New Zealand Speleological Society (unpublished). |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}